Category : NEET
Muscular Tissues
Contractility and motility (movement) are fundamental properties of protoplasm. That is why, all cells possess potential motility. Contraction for motility in the cells results essentially from the interaction of two contractile proteins, actin and myosin. These proteins enter into the composition of microfilaments of cellular cytoskeleton. During evolution, organism achieved enhancement in motility by various means. For example, unicellular organisms and cells of lower metazoans (mesozoa and parazoa) acquired the ability to form pseudopodia, flagella, or cilia as locomotory organelles. Then, in eumetazoans began the evolution of specialized contractile cells having much more of actin and myosin proteins. In the lowest eumetazoans (cnidarians), basal ends of epithelial cells are drawn out into elongated contractile processes. That is why, these cells are called epitheliomuscular or myoepithelial cells. In the eumetazoans higher than cnidarians, independent contractile cells, called muscle cells (myocytes), occur and form Muscular Tissues (muscles). These tissues are obviously responsible for movements of organs and locomotion of the body in response to stimuli. These develop from embryonic mesoderm except for those of the iris and ciliary body of eyes, which are ectodermal in origin. About 40% to 50% of our body mass is of muscles. The muscle cells are always elongated, slender and spindle-shaped, fibre-like cells, These are, therefore called muscle fibres. These possess large numbers of myofibrils formed of actin and myosin. The myofibrils are obviously stuffed in a small amount of cytoplasm due to thin, fibre-like shape of muscle cells. Muscle cells are highly contractile (contracting to 1/3 or 1/2 the resting length). Muscle cells lose capacity to divide, multiply and regenerate to a great extent. Study of muscle is called myology. Types of muscle are following -
(i) Striated or striped muscles : Most muscles of body are striated. These generally bring about voluntary movements under conscious control of brain and, hence, called voluntary muscles. Most of these are inserted at both ends upon bones in different parts of the body depend upon these muscles. Hence, these are also called skeletal muscles. Movements of limbs and the body solely depend upon these muscles. Hence these are also called somatic muscles. These are also called phasic type of muscles, because contraction in these is rapid, but brief and fatigue occurs quickly.
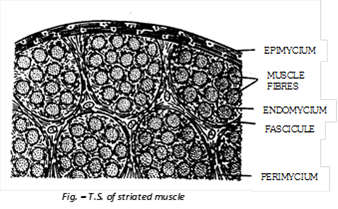
(a) Structure of striated muscles:
Each striated muscle consists of numerous muscle fibres segregated into several small and parallel bundles, called Fasciculi. Fibres of each fascicule are bound together by a connective tissue sheath, called endomycium. All fasciculi of a muscle are bound together by a connective tissue termed perimycium which also forms a sheath around each fascicule. Similarly, the whole muscle itself is covered by a connective tissue sheath, called epimycium. The latter extends as a tendons at each end of the muscle to insert it on to bones. Endomycium, perimycium and epimycium contain collagen and elastin fibres and also reticular fibres in some muscles.
(b) Structure of striated muscle fibres :
Striated muscle fibres are 0.01mm to 30 cm long, slender, cylindrical, unbranched (except some of the tongue and facial muscles) and multinucleated. Their covering envelope is called sarcolemma. The homogeneous, semifluid cytoplasm is called sarcoplasm. Two types of fibres can be recognized in most striated muscles, viz. white fibres and red fibres. Three soluble proteins myoalbumin, myoglobin (a haemoglobin like protein capable of binding oxygen) and myogen, a number of enzymes, glycogen granules and lipid droplets are found in the sarcoplasm of all fibres. The formed elements include many oval and compressed nuclei, mitochondria (sarcosomes), a small golgi apparatus, sarcoplasmic (endoplasmic) reticulum, etc. The red fibres are shorter, have more numerous and more deeply situated nuclei, more myoglobin and more numerous mitochondria producing more ATP. These are thus, adapted for prolonged and continued muscle activity required to support the body against gravity and for long continuing athletic events like marathon races. These are, therefore also called slow fibres. The white fibres (fast fibres) are longer, peripherally situated nuclei and fewer mitochondria, but these degrade glucose rapidly by glycolytic process to obtained energy at a faster rate. Thus, these are adapted for very rapid and powerful muscle contractions required for jumping, fast running etc. Due to multinucleate condition each striated muscle fibre is regarded a multicellular syncytial body, instead of a single cell. This is so, because each fibre is formed by fusion of a number of embryonic stem cells, called myoblasts. Most part of each striated muscle fibre is studded with several hundred to several thousand longitudinally oriented and parallel, slender myofibrils or sarcostyles.
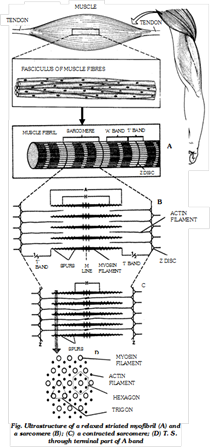
(c) Fine structure of striated muscle fibres : Striated muscle fibres shows transverse striation in the form of regular alternate dark A (anisotropic) and light I (isotropic) bands. The ‘A’ band contains about 120Å thick and 1.8 µ long “myosin filaments”. The I band contains about 60Å thick and 1.0µ long “actin filament” which are twice as many as myosin filaments. Each I band is divided into two equal halves by a thin, fibrous and transverse zig-zag partition, called ‘Z’ band (‘ Z’ disc) or Krause’s membrane. Each segment of a fibril between two adjacent ‘z’ bands is called a sarcomere. It is 2.3µ long in uncontracted mammalian striated fibres. A slender transverse line, the ‘M’ or Hansen’s line is visible in middle of each ‘A’ band. The major, middle region of ‘A’ band is comparatively lighter, but its terminal parts appear darker. The middle lighter region is called ‘H’ zone. Due to the geomatric bonding pattern, the end of each myosin filament is, thus, encircled by the ends of six actin filaments (hexagon), while the end of each actin filaments is encircled by the ends of three myosin filaments (trigon).
(d) Ultrastructure of myofilaments : At the molecular level, each myosin filament is composed of about 500 thread-like myosin molecules. Three different kinds of proteins participate in the composition of actin filaments. The major part of an actin filament is a coiled double helical strand whose each arm is a linear polymer of small and globular molecules (monomers) actin protein. Another coiled double helical, but thiner, strand runs along the whole length of actin strand. Each arm of this strand is a polymer of fibre-like molecules of tropomyosin protein. The third protein is troponin.
(e) Working of striated muscles : H.E. Huxley and A.F. Huxley in 1954 proposed a theory to explain the process of muscular contraction. This theory is known as sliding filament theory. It was observed that when a fibril contracts, its ‘A’ bands remain intact, while the ‘I’ bands progressively shorten and eventually disappear when the fibril has shortened to about 65% of its resting length. At this stage. ‘H’ zones also disappear because the actin filaments of both sides in each sarcomere reach, and may even overlap each other at the “M” line, and the ‘Z’ lines now touch the ends of myosin filaments. It was further observed that if a fibre is mechanically streched, the zones of overlap between thick and thin filaments are shorter than in resting condition, resulting in wider ‘H’ zones. These observations led Huxley to propose that shortening of the fibrils in contraction is brought about by sliding movement of actin filaments over myosin filaments towards “M” line by means of rapidly forming and breaking cross bridges or rachets at the spurs of myosin filaments. Thus, the sarcomere were recognised as the ‘ultimate units of contraction’.
It was also proved that the ‘A’ band occupies about 2/3 length of a sarcomere, so that as its maximum contraction, a muscle shortens by about 1/3 of its length. Actin has a strong affinity to myosin; if uninhibited, it readly combines with myosin to form a contractile complex called actomyosin. Normally, tropomyosin and troponin of actin filaments inhibit formation of actomyosin. As a muscle fibre is stimulated by a motor nerve fibre, a large number of calcium ion (Ca++) are released from the sarcoplasmic reticulum. Troponin has a strong affinity to (Ca++). As these ions combine with troponin, the tropomyosin troponin complex is inactivated so that actin is now free to interact with the spurs of corresponding myosin filaments. Thus cross bridges of actomyosin are formed at the spurs.
The energy required for this interaction is provided by hydrolysis of ATP molecules in the spurs of myosin which contain ATPase enzyme for this purpose. Immediately after this process the Ca++ are forcefully pumped back into the endoplasmic reticulum by spending a large amount of energy of ATP. This reactivates the tropomyosin - troponin complex which, in turn inactivates the actin. The actomyosin of cross bridges, therefore, split back into actin and myosin and spurs spring back to their normal position. As it clears from above account, muscle fibres requires a large amount of energy, not only for contraction, but also for relaxation (pumping back of Ca++ into endoplasmic reticulum). That is why, these fibres contain more numerous mitochondria and produce more ATP. Besides this, an additional “high-energy compound” creatine phosphate, acts as a “reservoir of high energy phosphate bonds” in these fibres.
(f) Stimulation of striated muscles : Striated muscles are neurogenic, i.e., these are stimulated to contract by somatic motor nerve fibres of central and peripheral nervous systems. A motor neuron innervates a group of muscle fibres by its axon terminalis, forming a motor fibres by its axon terminalis, forming a motor end plate. The motor neuron, together with the fibres it innervates, constitutes a “motor unit (neuromotor unit)”. The junctions of the axon terminalis with the muscle fibre are called neuromuscular or myoneural junctions. The axon terminals release acetylcholine at these junctions to transmit excitation impulses to the sarcolemma of the fibres. Acetylcholine depolarises the sarcolemma and thus triggers a self-propagating action potential spreading towards both ends of the fibres. The conduction of the impulse in the sarcolemma is electrochemically similar to that found in the neurons.
(ii) Smooth muscles : These are called smooth, plain nonstriated involuntary or unstriped muscles due to absence of striations. These occur in the walls of hollow internal organs (alimentary canal, gall bladder, bile ducts, respiratory tracts, uterus, urinogenital ducts, urinary bladder, blood vessels, etc.), in capsules of lymph glands, spleen etc., in iris and ciliary body of eyes, skin dermis, penis and other accessory genitalia, etc. There is no connection of these muscles with bones. Smooth muscles of skin dermis, called arrector pilli muscles, are associated with hair roots, and are responsible for goose-flesh (erection of hairs). Those of penis form a muscular network which helps in its erection and limping.

(a) Structure : Smooth muscle fibre is unbranched spindle shaped, uninucleated and has no sarcolemma. Contraction is slow, involuntary under the control of ANS. Functionally smooth muscles are of two types:
(1) Single-unit smooth muscle : Single unit smooth muscle fibres are composed of muscle fibres closely joined together, contract as a single unit. e.g., urinary bladder and gastrointestinal tract.
(2) Multi-unit smooth muscles : Are composed of more independent muscle fibres, contract as separate units e.g. - hair root muscle, muscles on the wall of large blood vessels, ciliary muscles and muscles of iris.
Contractility in smooth muscle fibres also depends upon interaction between myosin and actin filaments. At chemical and mechanical levels, the mechanism of contraction and relaxation in smooth muscle fibres is basically the same as in striated fibres. Smooth muscles are innervated only by the fibres of autonomic nervous system. The multiunit muscles are principally neurogenic, i.e., these contract on nervous stimulations. The visceral or single unit muscles are, on the other hand, principally myogenic, i.e., self-excitatory, because the action potential is generated spontaneously within the muscles themselves due to non-nervous intrinsic factors like mechanical stretching, temperature, or chemical stimulation by hormones and other substances. For example, when the gut wall gets stretched due to arrival of food, its muscles undergo rhythmic contractions and relaxations, bringing about the well-known peristalsis of gut wall. Similar peristalsis occurs in most other tubular viscera.
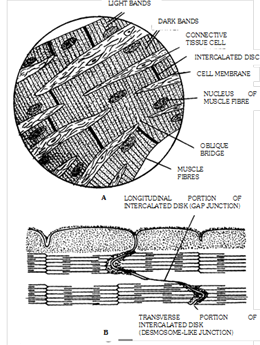
(iii) Cardiac muscles: Heart wall (also the wall of large veins just where these enter into the heart) is made up of cardiac muscles and, hence, called myocardium. Structurally, these muscles resemble striated muscles but, functioning independently of the conscious control of brain, these are involuntary like the smooth muscles. Cardiac muscle cells of fibres are comparatively shorter and thicker, cylindrical, mostly uninucleate with a central nucleus, somewhat branched and covered by a sarcolemma. The mechanism of contraction and relaxation in fibres of these muscles is the same as in skeletal muscle fibres, except that the duration of contraction is much longer in cardiac muscle fibres. Cardiac muscles require a large amount of energy of ATP for their incessant working throughout life. That is why, fibre of cardiac muscles contain comparatively very numerous, large and elongated mitochondria. Cardiac muscles cells divided at places by intercalated disc. Cardiac muscles are contract quickly and do not get fatigued. These muscles continue rhythmic contraction throughout life under the control of ANS. Heart is composed of three cardiac muscles viz. atrial muscle, ventricular muscle and self-excitatory and conducting muscle.
Differences between three types of muscle fibres
|
S.No. |
Feature |
Striated or Striped or Skeletal or Voluntary muscle fibres |
Non-striated or Unstriped or Smooth or Visceral or Involuntary muscle fibres |
Cardiac muscle fibres |
|
1. |
Shape |
Long cylindrical |
Fusiform (thick in middle tapering at ends) (0.02 nm to 0.2 nm long) |
Network of fibres |
|
2. |
Stripes |
Dark A bands and light I bands present |
Absent |
Present |
|
3. |
Nucleus |
Many (syncytial) at periphery |
Single at the centre of each cell |
Many nuclei between successive end plates central position |
|
4. |
Unit |
Sarcomeres, cylindrical long myofibrils placed end to end forming cylindrical myofibrils |
Fusiform cells with inconspicuous borders |
Oblique cross-connecting fibres make this muscle an interconnected bundle of myofibrils |
|
5. |
Attachment |
To bones |
To soft organs or viscera |
Not attached to other organs except major blood vessels which are isolated and covered by pericardium |
|
6. |
Sarcolemma |
Distinct |
Absent |
Absent |
|
7. |
Sarcoplasmic Reticulum |
Well developed |
Less extensive |
Poorly formed |
|
8. |
Blood supply |
Rich |
Poor |
Rich |
|
9. |
Contraction |
Quick, fatigue fast |
Slow, sustained contraction |
Rhythmic, contractions originate in heart (pace maker immune to fatigue) |
|
10. |
Location |
Generally peripheral, tongue, proximal part of oesophagus |
Central, in hollow visceral organs, iris of the eye, dermis of the skin |
Only in heart |
|
11. |
Intercalated discs |
Absent |
Absent |
Present |
|
12. |
T-tubule system |
Well developed |
Lacking |
Well developed |
|
13. |
Innervated nerves |
Motor nerves from central nervous system (neurogenic) |
Nerves from autonomic nervous system (neurogenic) |
Nerves from central and autonomic nervous system (myogenic) |
|
14. |
Fibres |
Unbranched |
Unbranched |
Fibres join by short oblique bridges |
|
15. |
Action |
Voluntary |
Involuntary |
Involuntary |
Important Tips
Poliomyelitis - Viral disease that weakens the muscles.



You need to login to perform this action.
You will be redirected in
3 sec

