Microsporogenesis
Category : NEET
Microsporogenesis
Microsporogenesis
The process of the formation and differentiation of microspores (pollen grains) from microspore mother cells (MMC) by reductional division is called microsporogenesis.
Microsporogenesis is well studied under following heads:
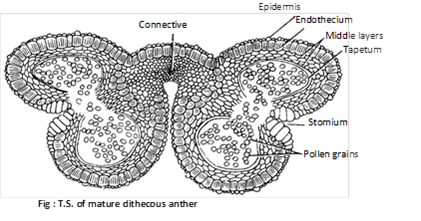
(1) Structure of anther: The fertile portion of stamens is called anther. Each anther is usually made up of two lobes connected by a connective. In turn each anther lobe contains two pollen chambers placed longitudinally. Each pollen chamber represents a microsporangium and is filled with a large number of pollen grains or microspores.
A typical anther consist of four microsporangia (tetrasporangiate) and such anthers is called dithecous e.g. mostly plants. In members of Malvaceae anthers are reniform or kidney shaped and consist of two microspoangia (bisporangiate), such anthers is called monothecous. In the smallest parasitic angiosperm, Arceuthobium minutissimum, anthers consist of only one microsporangium (monosporangiate).
The pollen sacs are surrounded by following 4 layers :
(i) Epidermis : This is the outermost single layered and protective. In Arceuthobium, cells of epidermis develops a fibrous thickening and the epidermis is designated as exothecium.
(ii) Endothecium : Inner to epidermis, there is a single layer of radially elongated cells. Cells of endothecium develop fibrous thickening (made up of cellulose with a little pectin and lignin) which help in the dehiscence of anther. In between these cells, a few cells without thickening are also present. These thick walled cells collectively form the stomium.
(iii) Middle layer : Three to four layers of thin walled cells situated just below the endothecium are known as middle layers. Cells of this layer are ephemeral and degenerate to provide nourishment to growing microspore mother cells.
(iv) Tapetum : This is the innermost layer of the wall. The cells are multinucleate (undergo endopolyploidy) and polyploid. Tapetal cells are nutritive.
In these cells the Ubisch bodies are present which help in the ornamentation of microspore walls. A compound sporopollenin is secreted in the exine of microspore wall. According to Periasamy and Swamy (1966), developmentally the tapetum has dual nature.
The tapetum is of two types :
(a) Amoeboid or Periplasmodial tapetum : In young condition cell wall of tapetal cells breaks, so protoplast of these cells become free between microspore mother cell and form mass of tapetal periplasmodium. e.g. Alisma, Typha, Tradescantia.
(b) Secretory or Glandular tapetum : This is the most common type of tapetum which remains insitu as such throughout. The tapetal cells secretes nourishment that passes into sporogenous cells. This tapetum attains its maximum development at the stage of pollen tetrads and then degenerates.
(2) Development of anther and formation of microspores (Pollen grains): The young anther consists of homogenous mass of paranchymatous cells surrounded by epidermis. It soon becomes four lobed. In each of the four lobes, some of the hypodermal cells begin to act as archesporial initials. Each archesporial initial divides into an outer primary parietal cell and an inner primary sporogenous cell. The primary parietal cell divides to form 3-5 wall layers, i.e., endothecium, middle layers and tapetum. The primary sporogenous cells divide to produce a mass of sporogenous cells or microsporocytes.
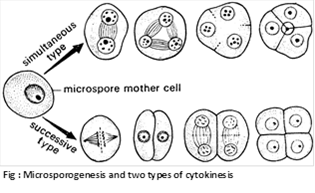
Each microspore mother cell divides meiotically to form four haploid microspores or pollen grains and remains arranged in tetrads. The arrangement of pollen grains in a tetrad is affected by cytokinesis during meiosis. It is of two types:
(i) Simultaneous type: The cytokinesis occurs simultaneously at the end of meiosis II to form tetrahedral tetrad. Here wall formation is centripetal. It is common in dicotyledons.
(ii) Successive type: The cytokinesis occurs twice i.e. each of the two nuclear division is followed by wall formation to form an isobilateral tetrad. Here the wall formation is centrifugal. It is found in monocotyledons.
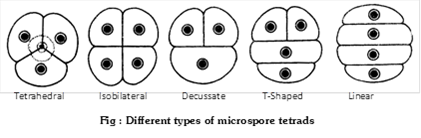
Besides tetrahedral and isobilateral tetrads, other types of tetrads are linear (e.g. Halophila), T-shaped (e.g. Aristolochia, Butomopsis) and decussate (e.g. Magnolia).
Now the microspores are separated from tetrad. In Drosera, Typha, Elodea, Hydrilla, etc. all the four pollen grains do not separate and thus form compound pollen grains. In the members of the family Cyperaceae (Cyprus), out of 4 pollen in a tetrad, 3 degenerate and one remains alive. So one meiosis produces one pollen. Sometimes more than four pollens are produced from one microspore mother cell. It is called as polyspory e.g. Cuscuta. In Calotropis (Asclepiadaceae) and some orchids all the pollen grains of an anther lobe form a typical structure called pollinium.
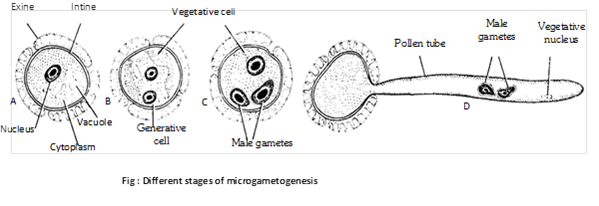
(3) Development of male gametophyte (Microgametogenesis): Microspore or pollen grain is the first cell of male gametophyte (partially developed). It is unicellular and haploid. The shape varies from oval to polyhedral. The wall of the pollen grain is made of two layers.
The outer layer is called exine. It is made up of sporopollenin (derived from carotenoid). It is thick and ornamented. At certain places, exine remains unthickened or missing and these places are known as germ pores. Sporopollenin is resistant to physical and biological decomposition. So pollen wall preserved for long periods in fossil deposits. The inner intine is thin, delicate and is made of cellulose and pectose.
In insect pollinated flowers, the exine of the pollen grain is covered with a yellowish, viscous and sticky substance called pollenkitt. This is perhaps the protective envelope which also sticks to the body of the insects and thus helps in pollination. It is chiefly made up of lipids and carotenoids. In monocots germ pores are absent and there is one germinal furrow. The development of male gametophyte from pollen grain is called microgametogenesis.
(4) Pre-pollination development: Microspores starts germinating in situ (i.e. while enclosed inside the microsporangium or pollen sac) and is called precocious. Microspores may be best defined as partially developed male gametophyte. Microspore nucleus divides mitotically to form a smaller generative cell lying next to spore wall and a much larger vegetative cell (or tube cell). A callose layer is deposited around the generative cell. The generative cell loses its contact with the wall of microspore and becomes free in the cytoplasm. The callose layer than dissolves. The pollen grains are shed from the anther at this bicelled stage (rarely three celled).
(5) Post-pollination development: The liberated pollen grains are transferred to the receptive surface of the carpel (i.e., stigma) by the process called pollination. On the stigma, the pollen grain absorbs water and swells within a few minutes. It releases the wall-held recognition factors. These factors determine whether the pollen grain will germinate on the stigma or not. Subsequent to mutual recognition, the vegetative (or tube) cell enlarges and comes out through one of the apertures in the form of a pollen tube. The wall of pollen tube is the extension of intine. The tube secretes exogenous pectinases and other hydrolytic enzymes to create a passage for its entry. It absorbs nourishment from the transmitting tissue of the style. Gradually, the vegetative and generative nuclei are carried by the pollen tube, the farmer lying at its tip. The generative cell divides to form two non-motile male gametes. The tube nucleus has no important function and may disintegrate.




You need to login to perform this action.
You will be redirected in
3 sec

