The physiology of blood circulation was first described by Sir William Harvey in 1628. The blood circulation in our body is divisible into 3 circuits -
(1) Coronary circulation : It involves blood supply to the heart wall and also drainage of the heart wall.
(i) Coronary arteries : One pair, arising from the aortic arch just above the semilunar valves. They break up into capillaries to supply oxygenated blood to the heart wall.
(ii) Coronary veins : Numerous, collecting deoxygenated blood from the heart wall and drains it into right auricle through coronary sinus which is formed by joining of most of the coronary veins. But some very fine coronary veins, called venae cordis minimae open directly in the right auricle by small sized openings called foramina of Thebesius.
(2) Pulmonary circulation : It includes circulation between heart and lungs. The right ventricle pumps deoxygenated blood into a single, thick vessel called pulmonary aorta which ascends upward and outside heart gets divided into longer, right and shorter, left pulmonary arteries running to the respective lungs where oxygenation of blood takes place. The oxygenated blood from lungs is returned to the left auricle by four pulmonary veins. Left auricle pumps this blood into the left ventricle.
(3) Systemic circulation : In this, circulation of blood occurs between heart and body organs. The left ventricle pumps the oxygenated blood into systemic arch which supplies it to the body organs other than lungs through a number of arteries. The deoxygenated blood from these organs is returned to the right auricle through two large veins (precaval and post caval). Right auricle pumps this blood into the right ventricle. Thus, the sytemic circulation involves two circuits -
Arterial system
It involves aorta, arteries, arterioles and meta-arterioles. It supplies oxygenated blood to all parts of the body except lungs.
The left ventricle of the heart pumps the oxygenated blood into a single, question marked shaped, long vessel called left carotid-systemic aorta. It is the largest blood vessel of the body.
After ascending from the heart, the systemic aotra turns and descends down to the level of lower border of fourth lumbar vertebra. At its distal extremity, it bifurcates into right and left common iliac arteries. The sytemic aorta has following parts -
(1) Ascending aorta : It gives off left and right coronary arteries.
Brachiocephalic (innominate) : Unpaired, largest branch of the aorta divides into right subclavian towards right side and right common carotid towards left side. Right subclavian gives off vertebral artery (supplies to head and part of right shoulder) and then enters into right arm, now called axillary artery or brachial artery, which divides into ulnar and radial arteries in the region of elbow. The right common carotid, enters into head and divides into external and internal carotids which supply the right parts of head by their branches.
Left common carotid : Unpaired artery, enters into head and divides into left external and internal carotids which supply the left parts of the head more...
Human respiratory system is derived from endoderm. Human respiratory system may be divided into two components-
(1) Respiratory tract or conducting portion
(2) Respiratory organs
(1) Respiratory tract or conducting portion : It is the passage for the air. In this part gaseous exchange does not takes place. It is also called dead air space. It is divided in following parts*-answer-*
(i) Nose (Latin-Nasa) (Greek-Rhine) : Cavity of nose is called nasal cavity. Nasal cavity is divided into two parts by nasal septum called mesethmoid. Each part is called nasal chamber. Each nasal chamber opens out side by external nares. Nasal septum has two part. First part is small and is made of cartilage (hyaline). Second part is major and it is bony. Vomer is the main bone. Each nasal chamber has three region.
(a) Vestibular region : Vestibular region also known as vestibule, lined by non keratinized squamous epithelium, it is ectodermal in origin and have sebaceous gland, sweat gland and hair. Vestibule is also found in inner air larynx, mouth and vagina. It acts like a seive to check the entry of large dust particles and other things.
(b) Respiratory region : Middle region lined by respiratory epithelium which is ciliated pseudostratified columnar epithelium. It contains mucus and serous cells. Mucus cells produce mucus and serous cells produce watery fluid. Respiratory epithelium is highly vascular and appears pink or reddish. Respiratory region acts as a air conditioner and makes the temperature of in going air nearly equal to body. It also acts as a filter not give entry to dust particles, flies or mosquitoes.
(c) Olfactory region : It is upper region. It is lined by olfactory epithelium. This is also called Schneiderian epithelium. Olfactory region is the organ of smell and detect the odour of inspired air. Inspiration is stopped if odour of air is foul or offensive. According to new researches pheromone receptors are found in nasal cavities.
(ii) Nasal conchae : Lateral wall of nasal cavity have three shelves like structures called conchae or turbinate. 3 pairs of nasal conchae are found. Nasal conchae are covered with mucus membrane. They increase the surface of nasal chamber. Both the chambers of nasal cavity open into nasopharynx by their apertures called internal nostrils or conchae. Adjacent to internal nostril there are opening of eustachian tube. Names of these three conchae and names of the bones that form them are given below.
(a) Superior conchae : The dorsal most chochae is supported mainly by nasal bone called nasoturbinate. It is the smallest conchae.
(b) Middle conchae : Ethmoid bone called ethmoturbinate.
(c) Inferior conchae : The ventral most conchae supported by maxilla bone called maxilloturbinate. It is a separate bone itself.
(iii) Pharynx : It is the short vertical about 12 cm long tube. The food and air passages cross here. It can be divided in 3 parts -
(a) Nasopharynx : Nasopharynx is only respiratory upper part in which internal nares open. There are 5 opening in more...
In men the respiratory organ are a pair of lung. Some snakes have unpaired lungs. Respiration by lungs is called pulmonary respiration. Lungs are found in all vertebrates except fishes. In Lung fishes such as protopterus, neoceratodus and lepidosiren air bladder is found, which is modified lung. Respiration in men and rabbit is pulmonary.
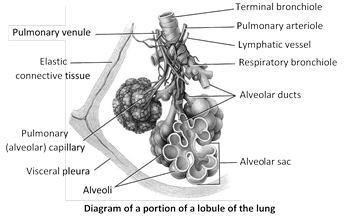
Lungs : Lungs lie in thoracic cavity on both side of heart in mediasternum space. Base of lung is attached to diaphragm. Right lung is divided into 3 lobes viz. Superior, Middle, Inferior and left lung is divided into two lobes Superior and Inferior. In rabbit, the left lung is divided into two lobes left anterior and left posterior where as the right lung has four lobes anterior azygous, right anterior, right posterior and posterior azygous. Lungs of reptiles are more complex than those of amphibians. In birds lungs are supplemented by elastic air sacs which increase respiratory efficiency. The narrow superior partion of lung is termed the apex or cupula.
Each lung is enclosed in two membrane called pleura. Pleura are layers of peritonium of thorax. Inner membrane is called the visceral pleuron. It is firmly bound to surface of lungs. The outer membrane is called parietal pleuron. It is attached to chest wall or wall of thoracic cavity. A narrow space exists between the two pleura. It is called pleural cavity. In pleural cavity a watery fluid is found called pleural fluid. Pleural fluid is glycoprotein in nature and secreted by pleura. Pleural fluid lubricate the pleura so that they may slide over each other without friction. This fluids reduces friction bewteen the membrane. When the lungs expand and contract in respiration. Pressure inside pleural cavity is negative - 5 mm Hg. Plurisy is inflamation of pleura and cause collection of fluid in pleural cavity. It results painful breathing (dyspnea). The surface of lung lying against the ribs, known as coastal surface. The mediastinal (medial) surface of each lung contains a region - the hilus, through which bronchi, pulmonary blood vessels, lymphatic vessels and nerve enter and exit.
Pulmonary volumes and capacities
The apparatus commonly used to measure the volume of air exchanged during breathing and the rate of ventilation is a spirometer (spiro=breathe) or respirometer. The record is called a spirogram. There are 4 respiratory volumes and capacity.
Respiratory volumes
(1) Tidal volume (TV) : Volume of air inspired or expired in relaxed or resting position – 500 ml. It consists of 150 ml of dead space volume and 350 ml of alveolar volume.
(2) Inspiratory reserve volume (IRV) : By taking a very deep breath, you can inspire a good deal more than 500 ml. This additional inhaled air, called IRV is about 3000 ml.
(3) Expiratory reserve volume (ERV) : If you inhale normally & then exhale as forcibly more...
(1) Skin : Respiration by skin is called cutaneous respiration. Skin is the only respiratory organ in most annelids (earthworm and leeches) and an additional respiratory organ in amphibians (Toads and frogs). Skin should be thin, moist, naked, permeable and well vascular for respiration. For cutaneous respiration animal should have large surface area then its volume and should have relatively inactive life to minimize the use of oxygen. Some marine annelids such as sandworms (nereis) have parapodia (locomotory appendages) for respiration. In frog 100% cutaneous respiration during hibernatin. In all marine snakes 20% respiration by skin.
(2) Tracheae : In insects, peripatus centipedes and millipedes tracheae are found for respiration. Tracheae are complex system of whitish, shining, intercommunicating air tubules. Tracheae are ectodermal air tubes. In cockroaches, three pairs of longitudinal tracheal trunks are present all along the length of body which are further connected with each other with the help of transverse branches. The main tracheae give off smaller tracheae whose branch repeatedly form a network of trachioles throughout the body. Trachea internally lined by chitinous cuticle called intima, which spirally thickened to form taenidae. Trachioles without taenidae, trachioles lined by trachein protein. From each tracheal trunk three branches come out. The dorsal branch is supplied to the dorsal muscles where as ventral one to nerve cord and ventral muscles and middle one to the alimentary canal.
(3) Book lungs and book gills : Spiders ticks, mites and scorpion (belongs to class arachnida) have book lungs for respiration. In scorpion 4 pairs of book lungs are present. A book lung is a chamber containing a series of thin vascular, parallel lamellae arranged like the pages of book. Book gills are found in marine king crab or horse shoe crab.
(4) Gills : Aquatic animals such as prawn, unio, fishes, sea stars and tadpoles respire by gills. Respiration by gills called bronchial respiration. Gills are of two types -
(i) External gills : External gills are found in arenicola (lug worm), larvae of certain insects e.g. damsel fly and some amphibians e.g. necturus, siren, proteus, frog tadpole first develop external gills which are replaced by internal gills later.
(ii) Internal gills : The internal gills may be phyllobranch (prawn), monopectionata (pila) eulamellibrach (unio), lamellibranch, fillibranch (pisces). In all fishes, gills are hemibranch or demibranch and holobranch. In gills, gill lamellae are found which have capillary network. Water is drawn into gills \[\to \] blood flowing in the capillaries of gill lamellae absorb oxygen from water and release \[C{{O}_{2}}\to \] water containing \[C{{O}_{2}}\] is thrown out from gills. The 80% of \[{{O}_{2}}\] of incoming water is absorbed.
Oxygen content of respiratory media
Frog is an amphibious animal i.e. they are live in water as well as on land hence according to their adaptations they posses different modes of respiration, which are as follows -
Frog is an amphibious animal i.e. they are live in water as well as on land hence according to their adaptations they posses different modes of respiration, which are as follows -
(1) Cutaneous respiration : By the skin. Under water, during hibernation frog respires by only skin. On land cutaneous respiration continues as usual. Thus cutaneous respiration take place always. By cutaneous respiration frog fulfill its 30% need of oxygen.
(2) Buccopharyngeal respiration : Like the skin, the mucosa of buccopharyngeal cavity in frog is also ideally adapted for gaseous exchange. Hence, while quietly floating upon water surface, and even when resting upon land, frogs respire by their buccopharyngeal cavity also. During this process, the mouth, gullet and glottis remain closed, but nares remain open.
(3) Pulmonary respiration : In frog, pulmonary respiration accounts for about 65% of the total \[{{O}_{2}}\,-\]intake. It particularly occurs when frogs lead an active life during rains and spring; either hopping upon land in search of food, or actively breeding in water.
Importasnt concept of respiration
(1) Respiratory quotient (R.Q.) : Respiratory quotient is the ratio of carbon dioxide output to oxygen usage during respiration. It is measured by Ganong’s respirometer.
\[\text{R}\text{.Q}\text{.}=\frac{\text{Volume of }C{{O}_{2}}\text{ formed}}{\text{Volume of }{{O}_{2}}\text{ utilized}}\]
High RQ
Low RQ
Due to fat deposition
Due to fever
Due to muscle exercise
During glycolysis
In low \[{{O}_{2}}\] environment
Due to oxidation of pyruvic acid.
When \[C{{O}_{2}}\] is fixed
When \[C{{O}_{2}}\] retain in tissue
In hibernating mammals
Due to acidosis
Due to alkalosis
Due to diabetes
In starvation
During gluconeogenesis
During glyconeogenesis
The volume of RQ depends upon the type of fuel substance being utilized for energy production.
RQ of various subtrates
Respiration is a process which involves intake of oxygen from environment and to deliver it to the cells. It include stepwise oxidation of food in cells with incoming oxygen, elimination of \[C{{O}_{2}}\] produced in oxidation, release of energy during oxidation and storing it in the form of ATP. It takes place in three basic steps -
(1) Pulmonary ventilation : The first process, pulmonary (pulmo = lung) ventilation, or breathing, is the inspiration (inflow) and expiration (outflow) of air between the atmosphere and the lungs.
(2) External (pulmonary) respiration : This is the exchange of gases between the air spaces of the lungs and blood in pulmonary capillaries. The blood gains \[{{O}_{2}}\] and loses \[C{{O}_{2}}.\]
(3) Internal (tissue) respiration : The exchange of gases between blood in systemic capillaries and tissue cells is known as internal (tissue) respiration. The blood loses \[{{O}_{2}}\] and gains \[C{{O}_{2}}.\] Within cells, the metabolic reactions that consume \[{{O}_{2}}\] and give off \[C{{O}_{2}}\]and give off \[C{{O}_{2}}\]during production of ATP are termed cellular respiration.
Respiratory surface
The surface at which exchange of gases (\[C{{O}_{2}}\] and \[{{O}_{2}}\]) takes place is called respiratory surface. Respiratory surface must be vascular and have enough area for gas exchange. For example - plasma membrane in protozoa, body wall (skin) in annelids, alveocapillary membrane in men.
Respiratory medium
Oxygen is dissolved in air and water. Thus water an air are source of oxygen for animals and called respiratory medium. Water and air are external respiratory medium. Inside the body an internal respiratory medium is also found. This internal respiratory medium is tissue fluid.
Types of respiration : It is of two types
(1) Aerobic respiration : It occurs in the presence of molecular oxygen. The oxygen completely oxidizes the food to carbon dioxide and water, releasing large amount of energy. The organisms showing aerobic respiration, are called aerobes. It is found in most of animals and plants. Aerobic respiration is of two main types direct and indirect.
\[\underset{\text{Glucose}}{\mathop{{{C}_{6}}{{H}_{12}}{{O}_{6}}}}\,+\underset{\text{oxygen}}{\mathop{6{{O}_{2}}}}\,\to \underset{\text{Carbon dioxide}}{\mathop{6C{{O}_{2}}}}\,+\underset{\text{Water}}{\mathop{6{{H}_{2}}O}}\,+\underset{\text{Energy}}{\mathop{2830\,kJ}}\,\]
(i) Direct respiration : It is the exchange of environmental oxygen with the carbon dioxide of the body cells without special respiratory organs and without the aid of blood. It is found in aerobic bacteria, protists, plants, sponges, coelenterates, flatworms, roundworms and most arthropods.
(ii) Indirect respiration : It involves special respiratory organs, such as skin, buccopharyngeal lining, gills and lungs, and needs the help of blood. The respiration in the skin, buccopharyngeal lining, gills and lungs is respectively called cutaneous buccopharyngeal, bronchial and pulmonary respiration. Cutaneous respiration takes place in annelids, some crustaceans, eel fish, amphibians and marine snakes. It occurs both in water and in air. Buccopharyngeal respiration is found in certain amphibians such as frog and toad. It occurs in the air. Branchial respiration is found in many annelids, most crustaceans and mollusks, some insect larvae, echinoderms, all fishes and some amphibians. It occurs in water only. Pulmonary respiration is found in snails, pila, some amphibians and in all reptiles, birds and mammals. It takes place in air only.
(2) more...
The process of respiration is completed in 4 steps –
(1) Breathing or ventilation
(2) Exchange of gases or External respiration
(3) Transport of gases
(4) Cellular respiration
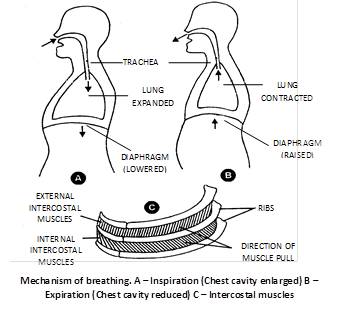
(1) Breathing or ventilation : Movement of thorax, expansion (inflation) and deflation of lungs and flow of air into the lungs and from the lungs. It is extracellular, energy consuming and physical process. Sum of inspiration and expiration is called respiratory movement. There are two steps of breathing -
(i) Inspiration : Intake of fresh air in lungs from outside. It is an active process. Blood pressure increases during later part of respiration. Following muscles are involve in inspiration.
(a) Diaphragm : Principle muscles of inspiration. Its skeletal muscles attached to sternum, vertebral column and ribs. It is formed by radial muscles fibres.
In relaxed condition it is dome shaped. Convex towards thoracic cavity and concave towards abdominal cavity. During inspiration it contract and become straight and descends down. This cause an increase in vertical diameter of thoracic cavity. Descent of diaphragm can explain about 75% of tidal air volume. 70% muscles fibres of diaphragm have some resistance to fatigue. Nerve which supply to diaphragm is phrenic nerve. Contribution of diaphragm in breathing of full term pregnant lady is 0%. Most important function of diaphragm of mammals is to aid in inspiration. If diaphragm is punctured, respiration will stop and patient will die.
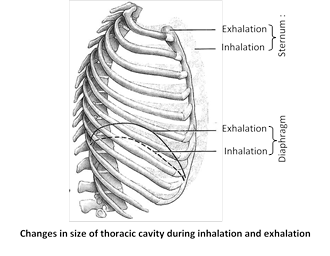
(b) External intercostal muscles : Gaps between the ribs are called intercostal spaces. They are filled by intercostal muscles. Intercostal muscles are of two types external intercostal muscles and internal intercostal muscles.
External intercostal muscles are related to inspiration and internal intercostal muscles are related to expiration. Here we are concerned with external intercostal muscles. External intercostal muscles start from lower border of upper rib and comes to end outer lip of upper border of lower rib. Thus direction of external intercostal muscles fibres is downward forward. Contraction of external intercostal muscles causes increase in anteroposterior diameter of thoracic cavity and transverse diameter of thoracic cavity.
This two dimensional increase in diameter (i.e. anteroposterior and transverse) of thoracic cavity is due to special arrangement of ribs. This increase of thoracic cavity assist by diaphragm the most important muscle of inspiration, it is dome-shaped skeletal muscle. Contraction of diaphragm causes it to flatten lowering its dome. For simplification we can assume that each rib attach anteriorly to sternum by its anterior end and posteriorly to vertebral column by its posterior end. Note these two points carefully -
Anterior end of rib is lower than the posterior end.
Middle portion of rib which is called shaft lies at lower level than the two end of rib (i.e. anterior and posterior)
(c) Accessory muscles of inspiration : These muscles normally are not called into action but more...
(1) Skin : Many aquatic animals, such as Hydra and starfish, excrete ammonia into the surrounding water by diffusion through the body wall. In land animals, the skin is often not permeable to water. This is an adaptation to prevent loss of body's water. Mammalian skin retains a minor excretory role by way of its sudoriferous, or sweat glands and sebaceous, or oil glands.
(i) Sweat gland : Sweat glands pass out sweat. The latter consists of water containing some inorganic salts (chiefly sodium chloride) and traces of urea and lactic acid. It also contains very small amounts of amino acids and glucose.
(ii) Sebaceous glands : Oil glands pass out sebum that contains some lipids such as waxes, sterols, other hydrocarbons and fatty acids.
(2) Lungs : Carbon dioxide and water are the waste products formed in respiration. Lungs remove the\[C{{O}_{2}}\]and some water as vapour in the expired air. Lungs have access to abundant oxygen and oxidise foreign substances, thus causing detoxification and also regulate temperature.
(3) Liver : Liver changes the decomposed haemoglobin of the worn-out red blood corpuscles into bile pigments, namely, bilirubin and biliverdin. These pigments pass into the alimentary canal with the bile for elimination in the faeces. The liver also excretes cholesterol, steroid hormones, certain vitamins and drugs via bile.
(4) Large intestine : Epithelial cells of the colon transfer some inorganic ions, such as calcium, magnesium and iron, from the blood into the cavity of the colon for removal with the faeces.
(5) Saliva : Heavy metals and drugs are excreted in the saliva.
(6) Gills : Gills remove \[C{{O}_{2}}\] in aquatic animals. They also excrete salt in many bony fish.
(1) Pyelonephritis : It is an inflammation of renal pelvis, calyces and interstitial tissue (G.pyelos = trough, tub; nephros = kidney; it is = inflammation). It is due to local bacterial infection. Bacteria reach here via urethra and ureter. Inflammation affects the countercurrent mechanism, and the victim fails to concentrate urine. Symptoms of the disease include pain in the back, and frequent and painful urination.
(2) Glomerulonephritis : It is the inflammation of glomeruli. It is caused by injury to the kidney, bacterial toxins, drug reaction, etc. Proteins and R.B.Cs pass into the filtrate.
(3) Cystitis : It is the inflammation of urinary bladder (G.kystis = bladder, - it is = inflammation). It is caused by bacterial infection. Patient has frequent, painful urination, often with burning sensation.
(4) Uremia : Uremia is the presence of an excessive amount of urea in the blood. It results from the decreased excretion of urea in the kidney tubules due to bacterial infection (nephritis) or some mechanical obstruction. urea poisons the cells at high concentration.
(5) Kidney stone (Renal calculus) : It is formed by precipitation of uric acid or oxalate. It blocks the kidney tubule. It causes severe pain (renal colic) in the back, spreading down to thighs. The stone may pass into the ureter or urinary bladder and may grow, and cause severe pain of blackade. When in bladder, the patient experiences frequent and painful urination and may pass blood in the urine. Surgery may be needed to remove stone and relieve pain.
(6) Kidney (Renal) failure (RF) : Partial or total inability of kidneys to carry on excretory and salt-water regulatory functions is called renal or kidney failure. Result kidney failure leads to (i) uremia, i.e., an excess of urea and other nitrogenous wastes in the blood (G.ouron = urine, haima-blood); (ii) Salt-water imbalance; and (iii) stoppage of erythropoietin secretion.
Causes : Many factors can cause kidney failure. Among these are tubular injury, infection, bacterial toxins, glomerulonephritis (inflammation of glomeruli) arterial or venous obstruction, fluid and electrolyte depletion, intrarenal precipitation of calcium and urates, drug reaction, heammorrhage, etc.
Artificial kidney
Artificial kidney, called haemodialyser, is a machine that is used to filter the blood of a person whose kidneys are damaged. The process is called haemodialysis. It may be defined as the separation of small molecules (crytalloids) from large molecules (colloids) in a solution by interposing a semipermeable membrane between the solution and water (dialyzing solution). It works on the principle of dialysis, i.e. diffusion of small solute molecules through a semipermeable membrane (G. dia = = through, lyo = separate). Haemodialyser is a cellophane tube suspended in a salt-water solution of the same composition as the normal blood plasma, except that no urea is present. Blood of the patient is pumped from one of the arteries into the cellophane tube after cooling it to 0oC and mixing with an anticoagulant (heparin). Pores of the cellophane tube allow urea, uric acid, creatinine, excess salts and excess H+ ions to diffuse from more...
(1) Protozoans : In protozoans like Amoeba and Paramecium carbon dioxide and ammonia are mostly excreted out by diffusion through general body surface. It is considered that the contractile vacuoles also play some role in the removal of excretory products.
(2) Sponges : In sponges, the nitrogenous metabolic waste (ammonia) leaves the body in the outgoing water current by diffusion.
Most of the sponges are marine and have no problem of surplus water in their cells. A few sponges lie in hypotonic fresh water and have contractile vacuoles in most of their cells.
(3) Coelenterates : Hydra also lacks special excretory organs. The nitrogenous waste products like ammonia are removed through the general surface of the body by diffusion. Some nitrogenous waste products are also thrown along with indigestible matter through the mouth.
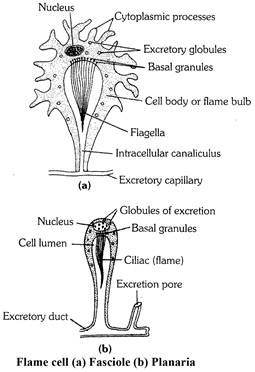
(4) Platyhelminthes : Planaria, liverfluke and tapeworm possess a large number of excretory cells called the flame cells (solenocytes) or protonephridia and long excretory ducts (also called canals of vessels). The flame cells open into the ductules which in turn open into the excretory duct.
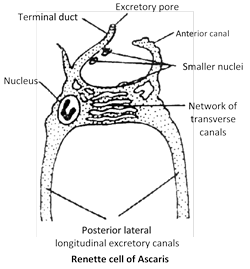
(5) Nematoda : The round worms such as Ascaris have H-shaped excretory system. It is made up of a single Renette cell. in entire length of body. It consists of two longitudinal excretory canals connected anteriorly by a network of transverse canals. A short terminal duct opens outside via excretory pore. Ascaris is excretes both ammonia and urea.
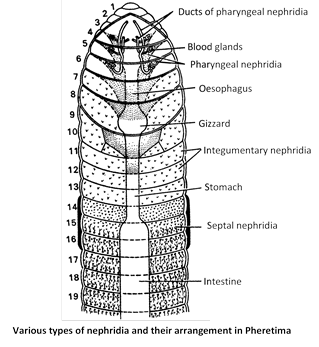
(6) Annelids (Earthworm) : In earthworm excretory system also known as nephridial system, which consist of 3 types of nephridia Pharyngeal, septal, and integumentary nephridia. All nephridia commonly called micronephridia. Earthworm excrete 40% urea, 20% ammonia, 40% amino acids. Earthworm mainly ureotelic. Chloragogen cells found in coelomic fluid also excretory in nature. Blood gland in earthworm found in 4, 5, 6 segment, serves for excretion, manufacture of blood corpuscles and Hb.
(7) Arthropods : The excretory system of the adult Prawn (crustacean) consists of a pair of antennary or green glands, a pair of lateral ducts and a single renal sac.
Insects, centipedes, millipedes and arachnids like scorpion and spider posses Malpighian tubules as their principal excretory organs. In the Malpighian tubules bicarbonates of potassium and sodium, water and uric acid are formed. A large amount of water and bicarbonates of potassium and sodium are reabsorbed by the cells of Malpighian tubules and then transferred to the blood (haemolymph). Uric acid is carried to the alimentary canal of the insect and is finally passed out through anus. Spiders and scorpions possess Malpighian tubules and coxal glands both for excretion.
more...
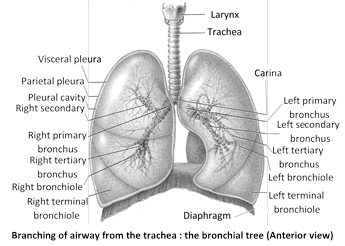 Each lung is enclosed in two membrane called pleura. Pleura are layers of peritonium of thorax. Inner membrane is called the visceral pleuron. It is firmly bound to surface of lungs. The outer membrane is called parietal pleuron. It is attached to chest wall or wall of thoracic cavity. A narrow space exists between the two pleura. It is called pleural cavity. In pleural cavity a watery fluid is found called pleural fluid. Pleural fluid is glycoprotein in nature and secreted by pleura. Pleural fluid lubricate the pleura so that they may slide over each other without friction. This fluids reduces friction bewteen the membrane. When the lungs expand and contract in respiration. Pressure inside pleural cavity is negative - 5 mm Hg. Plurisy is inflamation of pleura and cause collection of fluid in pleural cavity. It results painful breathing (dyspnea). The surface of lung lying against the ribs, known as coastal surface. The mediastinal (medial) surface of each lung contains a region - the hilus, through which bronchi, pulmonary blood vessels, lymphatic vessels and nerve enter and exit.
Each lung is enclosed in two membrane called pleura. Pleura are layers of peritonium of thorax. Inner membrane is called the visceral pleuron. It is firmly bound to surface of lungs. The outer membrane is called parietal pleuron. It is attached to chest wall or wall of thoracic cavity. A narrow space exists between the two pleura. It is called pleural cavity. In pleural cavity a watery fluid is found called pleural fluid. Pleural fluid is glycoprotein in nature and secreted by pleura. Pleural fluid lubricate the pleura so that they may slide over each other without friction. This fluids reduces friction bewteen the membrane. When the lungs expand and contract in respiration. Pressure inside pleural cavity is negative - 5 mm Hg. Plurisy is inflamation of pleura and cause collection of fluid in pleural cavity. It results painful breathing (dyspnea). The surface of lung lying against the ribs, known as coastal surface. The mediastinal (medial) surface of each lung contains a region - the hilus, through which bronchi, pulmonary blood vessels, lymphatic vessels and nerve enter and exit.
 Pulmonary volumes and capacities
The apparatus commonly used to measure the volume of air exchanged during breathing and the rate of ventilation is a spirometer (spiro=breathe) or respirometer. The record is called a spirogram. There are 4 respiratory volumes and capacity.
Respiratory volumes
(1) Tidal volume (TV) : Volume of air inspired or expired in relaxed or resting position – 500 ml. It consists of 150 ml of dead space volume and 350 ml of alveolar volume.
(2) Inspiratory reserve volume (IRV) : By taking a very deep breath, you can inspire a good deal more than 500 ml. This additional inhaled air, called IRV is about 3000 ml.
(3) Expiratory reserve volume (ERV) : If you inhale normally & then exhale as forcibly more...
Pulmonary volumes and capacities
The apparatus commonly used to measure the volume of air exchanged during breathing and the rate of ventilation is a spirometer (spiro=breathe) or respirometer. The record is called a spirogram. There are 4 respiratory volumes and capacity.
Respiratory volumes
(1) Tidal volume (TV) : Volume of air inspired or expired in relaxed or resting position – 500 ml. It consists of 150 ml of dead space volume and 350 ml of alveolar volume.
(2) Inspiratory reserve volume (IRV) : By taking a very deep breath, you can inspire a good deal more than 500 ml. This additional inhaled air, called IRV is about 3000 ml.
(3) Expiratory reserve volume (ERV) : If you inhale normally & then exhale as forcibly more... 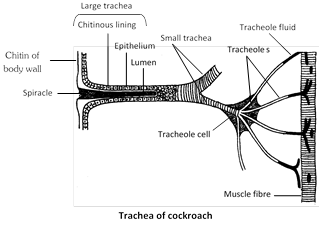 (3) Book lungs and book gills : Spiders ticks, mites and scorpion (belongs to class arachnida) have book lungs for respiration. In scorpion 4 pairs of book lungs are present. A book lung is a chamber containing a series of thin vascular, parallel lamellae arranged like the pages of book. Book gills are found in marine king crab or horse shoe crab.
(4) Gills : Aquatic animals such as prawn, unio, fishes, sea stars and tadpoles respire by gills. Respiration by gills called bronchial respiration. Gills are of two types -
(i) External gills : External gills are found in arenicola (lug worm), larvae of certain insects e.g. damsel fly and some amphibians e.g. necturus, siren, proteus, frog tadpole first develop external gills which are replaced by internal gills later.
(ii) Internal gills : The internal gills may be phyllobranch (prawn), monopectionata (pila) eulamellibrach (unio), lamellibranch, fillibranch (pisces). In all fishes, gills are hemibranch or demibranch and holobranch. In gills, gill lamellae are found which have capillary network. Water is drawn into gills \[\to \] blood flowing in the capillaries of gill lamellae absorb oxygen from water and release \[C{{O}_{2}}\to \] water containing \[C{{O}_{2}}\] is thrown out from gills. The 80% of \[{{O}_{2}}\] of incoming water is absorbed.
Oxygen content of respiratory media
(3) Book lungs and book gills : Spiders ticks, mites and scorpion (belongs to class arachnida) have book lungs for respiration. In scorpion 4 pairs of book lungs are present. A book lung is a chamber containing a series of thin vascular, parallel lamellae arranged like the pages of book. Book gills are found in marine king crab or horse shoe crab.
(4) Gills : Aquatic animals such as prawn, unio, fishes, sea stars and tadpoles respire by gills. Respiration by gills called bronchial respiration. Gills are of two types -
(i) External gills : External gills are found in arenicola (lug worm), larvae of certain insects e.g. damsel fly and some amphibians e.g. necturus, siren, proteus, frog tadpole first develop external gills which are replaced by internal gills later.
(ii) Internal gills : The internal gills may be phyllobranch (prawn), monopectionata (pila) eulamellibrach (unio), lamellibranch, fillibranch (pisces). In all fishes, gills are hemibranch or demibranch and holobranch. In gills, gill lamellae are found which have capillary network. Water is drawn into gills \[\to \] blood flowing in the capillaries of gill lamellae absorb oxygen from water and release \[C{{O}_{2}}\to \] water containing \[C{{O}_{2}}\] is thrown out from gills. The 80% of \[{{O}_{2}}\] of incoming water is absorbed.
Oxygen content of respiratory media


 (b) External intercostal muscles : Gaps between the ribs are called intercostal spaces. They are filled by intercostal muscles. Intercostal muscles are of two types external intercostal muscles and internal intercostal muscles.
External intercostal muscles are related to inspiration and internal intercostal muscles are related to expiration. Here we are concerned with external intercostal muscles. External intercostal muscles start from lower border of upper rib and comes to end outer lip of upper border of lower rib. Thus direction of external intercostal muscles fibres is downward forward. Contraction of external intercostal muscles causes increase in anteroposterior diameter of thoracic cavity and transverse diameter of thoracic cavity.
This two dimensional increase in diameter (i.e. anteroposterior and transverse) of thoracic cavity is due to special arrangement of ribs. This increase of thoracic cavity assist by diaphragm the most important muscle of inspiration, it is dome-shaped skeletal muscle. Contraction of diaphragm causes it to flatten lowering its dome. For simplification we can assume that each rib attach anteriorly to sternum by its anterior end and posteriorly to vertebral column by its posterior end. Note these two points carefully -
(b) External intercostal muscles : Gaps between the ribs are called intercostal spaces. They are filled by intercostal muscles. Intercostal muscles are of two types external intercostal muscles and internal intercostal muscles.
External intercostal muscles are related to inspiration and internal intercostal muscles are related to expiration. Here we are concerned with external intercostal muscles. External intercostal muscles start from lower border of upper rib and comes to end outer lip of upper border of lower rib. Thus direction of external intercostal muscles fibres is downward forward. Contraction of external intercostal muscles causes increase in anteroposterior diameter of thoracic cavity and transverse diameter of thoracic cavity.
This two dimensional increase in diameter (i.e. anteroposterior and transverse) of thoracic cavity is due to special arrangement of ribs. This increase of thoracic cavity assist by diaphragm the most important muscle of inspiration, it is dome-shaped skeletal muscle. Contraction of diaphragm causes it to flatten lowering its dome. For simplification we can assume that each rib attach anteriorly to sternum by its anterior end and posteriorly to vertebral column by its posterior end. Note these two points carefully -
 (c) Accessory muscles of inspiration : These muscles normally are not called into action but
(c) Accessory muscles of inspiration : These muscles normally are not called into action but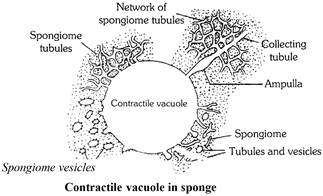 (3) Coelenterates : Hydra also lacks special excretory organs. The nitrogenous waste products like ammonia are removed through the general surface of the body by diffusion. Some nitrogenous waste products are also thrown along with indigestible matter through the mouth.
(4) Platyhelminthes : Planaria, liverfluke and tapeworm possess a large number of excretory cells called the flame cells (solenocytes) or protonephridia and long excretory ducts (also called canals of vessels). The flame cells open into the ductules which in turn open into the excretory duct.
(3) Coelenterates : Hydra also lacks special excretory organs. The nitrogenous waste products like ammonia are removed through the general surface of the body by diffusion. Some nitrogenous waste products are also thrown along with indigestible matter through the mouth.
(4) Platyhelminthes : Planaria, liverfluke and tapeworm possess a large number of excretory cells called the flame cells (solenocytes) or protonephridia and long excretory ducts (also called canals of vessels). The flame cells open into the ductules which in turn open into the excretory duct.
 (5) Nematoda : The round worms such as Ascaris have H-shaped excretory system. It is made up of a single Renette cell. in entire length of body. It consists of two longitudinal excretory canals connected anteriorly by a network of transverse canals. A short terminal duct opens outside via excretory pore. Ascaris is excretes both ammonia and urea.
(5) Nematoda : The round worms such as Ascaris have H-shaped excretory system. It is made up of a single Renette cell. in entire length of body. It consists of two longitudinal excretory canals connected anteriorly by a network of transverse canals. A short terminal duct opens outside via excretory pore. Ascaris is excretes both ammonia and urea.
 (6) Annelids (Earthworm) : In earthworm excretory system also known as nephridial system, which consist of 3 types of nephridia Pharyngeal, septal, and integumentary nephridia. All nephridia commonly called micronephridia. Earthworm excrete 40% urea, 20% ammonia, 40% amino acids. Earthworm mainly ureotelic. Chloragogen cells found in coelomic fluid also excretory in nature. Blood gland in earthworm found in 4, 5, 6 segment, serves for excretion, manufacture of blood corpuscles and Hb.
(6) Annelids (Earthworm) : In earthworm excretory system also known as nephridial system, which consist of 3 types of nephridia Pharyngeal, septal, and integumentary nephridia. All nephridia commonly called micronephridia. Earthworm excrete 40% urea, 20% ammonia, 40% amino acids. Earthworm mainly ureotelic. Chloragogen cells found in coelomic fluid also excretory in nature. Blood gland in earthworm found in 4, 5, 6 segment, serves for excretion, manufacture of blood corpuscles and Hb.
 (7) Arthropods : The excretory system of the adult Prawn (crustacean) consists of a pair of antennary or green glands, a pair of lateral ducts and a single renal sac.
Insects, centipedes, millipedes and arachnids like scorpion and spider posses Malpighian tubules as their principal excretory organs. In the Malpighian tubules bicarbonates of potassium and sodium, water and uric acid are formed. A large amount of water and bicarbonates of potassium and sodium are reabsorbed by the cells of Malpighian tubules and then transferred to the blood (haemolymph). Uric acid is carried to the alimentary canal of the insect and is finally passed out through anus. Spiders and scorpions possess Malpighian tubules and coxal glands both for excretion.
(7) Arthropods : The excretory system of the adult Prawn (crustacean) consists of a pair of antennary or green glands, a pair of lateral ducts and a single renal sac.
Insects, centipedes, millipedes and arachnids like scorpion and spider posses Malpighian tubules as their principal excretory organs. In the Malpighian tubules bicarbonates of potassium and sodium, water and uric acid are formed. A large amount of water and bicarbonates of potassium and sodium are reabsorbed by the cells of Malpighian tubules and then transferred to the blood (haemolymph). Uric acid is carried to the alimentary canal of the insect and is finally passed out through anus. Spiders and scorpions possess Malpighian tubules and coxal glands both for excretion.